1.1 纤维素晶型
天然的纤维素只存在一种纤维素晶型结构,即纤维素Ⅰ[9]。通过核磁共振碳谱(CP/MAS 13C NMR)研究发现,在细菌及藻类纤维素中,纤维素链主要堆砌排列成三斜晶体,定义为纤维素Ⅰα; 而高等植物(麻类、棉花、木材等)中的纤维素为单斜晶体结构,定义为纤维素Ⅰβ[10]。木质纤维原料中的纤维素Ⅰβ分子链平行排列,分别处于单斜晶体的四周和中心。葡萄糖分子的O6基团主要偏向C4,形成tg构象。在同一晶胞内,O3与相邻葡萄糖的O5形成分子内氢键,O2与O6之间的氢原子可形成多种形式的分子间氢键和分子内氢键。
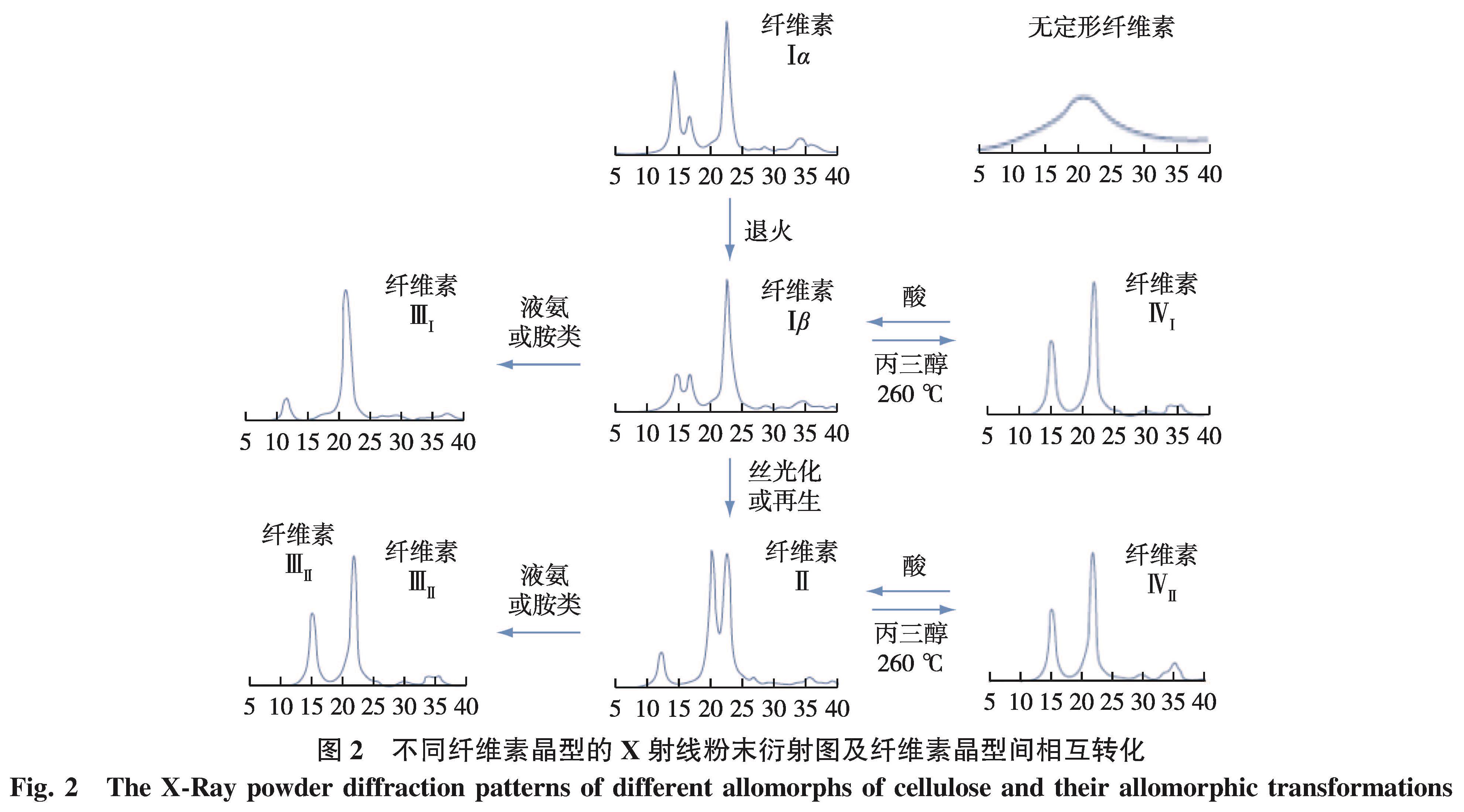
由于纤维素Ⅰβ中纤维素大分子的层状二维排列与致密氢键作用体系,该晶型在化学预处理和纤维素酶解过程中更难被解离。因此,纤维素晶型由稳定的纤维素Ⅰβ向其他晶型的转变逐渐成为提高木质纤维素酶解效率的有效措施之一。图2列举了已知的纤维素晶型转变所需条件及相应结构的X射线衍射(XRD)一维图谱。碱性蒸煮是最为常用的预处理方法之一,该法对木质纤维原料中纤维素及其他组分的结构均产生影响,同时也是改变纤维素晶型的经典方法[11]。笔者前期曾研究了梯度碱处理前后毛竹综纤维素的化学组成与纤维素聚集态结构变化[12]。结果表明,不同浓度的碱处理造成纤维素晶型由纤维素Ⅰ型(分子链平行排列)向纤维素Ⅱ型(分子链反平行排列)的递进转化,该变化伴随纤维素结晶区的减少、微纤丝润胀和宏纤丝束的剥离,有利于后续酶解时酶的渗入[13]。该结论与诸多报道中纤维素Ⅱ酶解效率变化趋势一致[14-17]。Jin等[18]将纤维素Ⅰ、Ⅱ型的微晶纤维素制备成去除无定型区的纳米晶体,观察其在石英晶体微天平(QCM-D)上的原子力显微镜(AFM)原位成像,发现两种纤维素晶型的晶粒形态差异较大,纤维素Ⅱ晶体呈颗粒状排列,具有更松散的聚集和更高的比表面积,有利于纤维素酶的吸附。纤维素Ⅱ的分子链呈反平行排列,同时存在角链与相邻内部分子链层附加的分子间氢键,不同于仅有层内氢键纤维素Ⅰ[19]。纤维素Ⅱ的角链存在O3—H…O5 分子内氢键,以及O6—H…O3的分子间氢键,氢键的平均长度为2.72 Å,比纤维素Ⅰ中的2.80 Å 稍短。因此纤维素Ⅱ比纤维素Ⅰ的热稳定性更好,同时也证明了纤维素Ⅰ向纤维素Ⅱ的转变是不可逆过程。
图2 不同纤维素晶型的X射线粉末衍射图及纤维素晶型间相互转化
Fig.2 The X-Ray powder diffraction patterns of different allomorphs of cellulose and their allomorphic transformations
研究表明,一些有机溶剂(如N-甲基吗啉-N-氧化物、氯化锂/二甲基乙酰胺、磷酸、离子液体等)对纤维素有一定的溶解能力,预处理后的木质纤维细胞壁发生明显的润胀解离[20-23]。在溶解后的体系中添加反向溶剂(如水、乙醇、甲醇等)后,纤维素析出再生,再生纤维素的晶型为纤维素 Ⅱ,其形貌也发生明显变化。此外,纤维素晶型转变过程中,分子间氢键的断裂重排,造成一部分结晶区的结构破坏,结晶度下降,孔隙增多[24]。Cheng等[25]研究了不同的木质纤维原料以1-乙基-3-甲基咪唑醋酸盐为溶剂分别在120和160 ℃ 预处理后的纤维素结晶结构变化。XRD和小角中子散射(SANS)表征证明了离子液体处理后的纤维素均发生由纤维素 Ⅰ 到纤维素Ⅱ的晶型转变,且在更高温度条件下处理时,晶型变化更为明显,酶解效率也更优异。因此,低结晶度的纤维素Ⅱ结构有利于酶解反应,再生纤维素的酶解效率相比存在于天然植物原料中的纤维素Ⅰβ大大提高[26]。但在不同的纤维素溶解体系中,再生纤维素Ⅱ的结晶度也有所差异,酶解效率不尽相同,其具体变化规律有待深入研究。
除天然纤维素Ⅰ和碱处理或溶解再生法得到的纤维素Ⅱ外,通过液氨爆破处理或者有机胺处理纤维素样品得到的纤维素Ⅲ也被深入研究。根据初始纤维素的晶型不同(纤维素Ⅰ或Ⅱ),得到的纤维素样品分别为纤维素ⅢⅠ和纤维素ⅢⅡ。这两种纤维素Ⅲ样品的衍射图样十分相似,但分子链的平行或反平行排列方式与其原始纤维素相同。纤维素Ⅲ中分子发生重排,O6基团偏向O5,形成tg构象。该构象使得纤维素Ⅲ分子内氢键数量减少,同时分子间氢键数量增多(图3)[27-28]。这类分子重排使更多可水解的葡聚糖链疏水平面暴露在表面,显著打破了纤维素的有序结晶结构造成的天然抗降解屏障,使酶解效率提升至原来的5倍。综上,纤维素的晶型转变通常伴随着分子链的重排,进而造成结晶度、结晶尺寸、晶面间距等结晶参数的变化。重排后的纤维素晶体一般具有更疏松的结构,有利于纤维素酶的反应,而这类结晶参数的转变也逐渐成为评价纤维素预处理效果的新指标。
图3 液氨处理纤维素Ⅰ转化为纤维素ⅢⅠ提高酶解效率示意图[27]
Fig.3 NH3 pretreatment transformation of cellulose Ⅰ to cellulose ⅢⅠ and enhancement of enzymatic hydrolysis performance
1.2 纤维素结晶度
纤维素具有由结晶区和非结晶区组成的两相结构,分子链排列规则的区域,即结晶区在纤维素中占的比例被定义为结晶度。纤维素的高结晶度是形成生物质抗降解屏障、阻碍酶解的主要因素之一。在木质纤维素的研究中,测定纤维素结晶度的方法很多,主要有XRD、红外光谱(IR)、NMR、拉曼光谱(RM)等,其中XRD为最常用的测定手段[29-32]。利用XRD图谱计算纤维素结晶度的方法分为3种:1)Segal法——通过结晶区峰高(I200-Iam)与200面的峰高(I200)的比值来计算结晶度。该方法运用范围最广,但已被证明其准确性有所欠缺[33-35]。2)结构精修法——借助数据处理软件,导入标准纤维素晶型,对实验所得XRD谱图进行分峰拟合,将代表结晶区纤维素的峰进行积分,并计算结晶区域峰强度与各衍射峰积分总强度的比值[36]。此方法相对准确性更高,逐渐得到广泛运用。3)将球磨纤维素作为无定形区的标样,衍射峰积分总强度减去无定形区,即得到结晶区域的衍射峰强度,从而计算出结晶度数值。
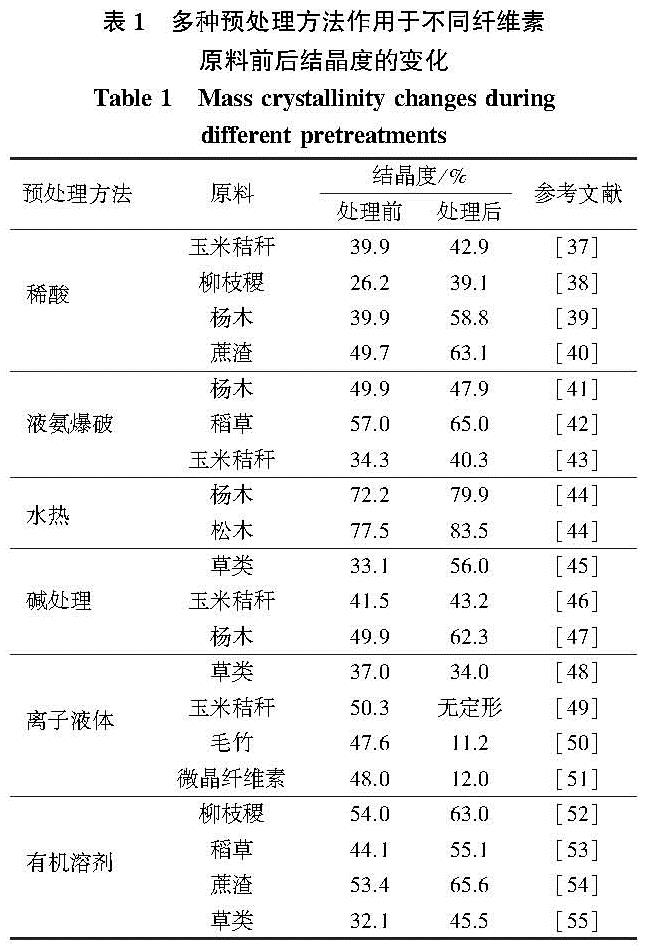
木质纤维素预处理过程中,结晶度变化显著。纤维素结晶度的降低,促使纤维素酶进入纤丝聚集体内核,吸附位点增多,从而提高了酶解效率,但并非所有的预处理均能达到理想的去结晶效果。表1总结了多种预处理方法作用于不同木质纤维原料前后结晶度的变化[37-55]。由表1可以发现,木质纤维素的结晶度变化趋势各异,主要随原料种类、预处理方法、反应条件等诸多因素而变化。对于常用的酸、碱、有机溶剂、离子盐等预处理,其主要目的是降解细胞壁中无定形的半纤维素、木质素,使包覆在纤维素周围的屏障去除,同时使微纤丝润胀,孔隙率和比表面积增加[56-57]。但是,这一过程通常伴随排列杂乱的无定形纤维素的降解,使得预处理物料中结晶纤维素的相对含量增加,结晶度反而更高,依然造成较高的生物质抗降解屏障。预处理强度进一步增加,如增加酸碱浓度、提高反应温度等,可更大程度地促进微纤丝的润胀并加剧纤维素链的重排。此时,纤维素结晶结构也随之打破,结晶度降低[58-59]。
降低纤维素结晶度的有效方式有机械处理法和溶解-再生法[60]。机械处理法,如球磨、超声、研磨法等操作简便,可迅速打破纤维素的高聚集态结构,降低分子量、结晶度等。笔者曾系统研究了球磨处理高结晶棉纤维的降解机制,发现球磨处理2 h后,纤维素结晶度从90%降至10%左右,同时分子量、粒径减小,比表面积和孔隙率增加,有利于纤维素酶的作用[32]。机械处理法常与化学预处理同时或分步进行,以实现木质纤维素的高效转化[61]。化学预处理可有效降解植物细胞壁中的木质素和半纤维素,实现组分分离,暴露纤维素微纤丝; 机械处理则有效降低物料的结晶度,两种处理方法共同作用,从多角度破解木质纤维素的抗降解屏障,促进生物转化效率的提升[62]。除机械处理外,通过溶解-再生法制得的纤维素具有较低结晶度,且其晶型为纤维素Ⅱ。Guo等[63]制备了低结晶度纤维素薄膜,并研究其酶解性能,发现即使薄膜中存在木质素,低结晶度样品的酶解速率更高。Arantes等[64]提出一种“无定形化”的纤维素酶解模型,认为纤维素酶解的过程可分为三步:纤维素酶吸附在纤维素的致密表面,此时纤维素分子链逐渐疏松,出现无定形化现象; 纤维素链在内切、外切葡聚糖酶的协同作用下被迅速水解为纤维二糖; 纤维二糖被β-葡萄糖苷酶进一步水解为葡萄糖。这种模型虽然得到一部分学者的认同,但对于低结晶度纤维素来说,不同方法制备的无定型区结构依旧存在差异,与纤维素酶的吸附、反应情况仍值得深入探索。
表1 多种预处理方法作用于不同纤维素原料前后结晶度的变化
Table 1 Mass crystallinity changes during different pretreatments
1.3 纤维素结晶尺寸与晶面间距
纤维素晶粒的大小(结晶尺寸)和晶面间距是衡量纤维素晶体性质的一项主要指标,对木质纤维素酶解糖化也具有较大影响。纤维素晶粒尺寸指微纤丝中相距最远的晶面间的宽度,与微纤丝中的分子链数量有关。普遍观点认为,单根基元纤维中含有36条纤维素分子链,呈六边形排列,但根据原料的不同,基元纤维可进一步拆分为18条或24条纤维素分子链[65]。一般来说,禾本科原料基元纤维中的纤维素链数量多于木、竹材原料[66]。在针叶木原料脱木质素过程的研究中,氧化脱木质素和亚氯酸钠脱木质素的处理条件不同对结晶度和结晶尺寸均产生影响[67]。氧化脱木质素和亚氯酸钠脱木质素都增加了针叶木中纤维素结晶区所占比例,且氧化脱木质素对结晶尺寸的影响更为明显。同时,结晶尺寸也随处理条件而变化。处理时间、浓度和处理温度等处理强度的提升使纤维素中晶粒聚集、结晶尺寸增加。结晶尺寸的大小决定了结晶区纤维素链的暴露程度,影响其与纤维素酶的接触。Nishiyama等[68]对杨木样品进行液氨爆破、蒸汽及稀酸预处理,结果表明3种预处理方法均提高了结晶尺寸。其原因可能是相邻的纤维素分子链在共晶面处发生了局部共结晶作用。液氨爆破预处理后杨木中纤维素晶型转化为纤维素Ⅲ,且结晶尺寸由未处理样品的2.4 nm增加至2倍以上(5.8 nm),远超过稀酸处理和蒸汽预处理后的结晶尺寸(3.1和4.1 nm),同时,代表200晶面的峰位(约2θ=22.5°)发生右移。根据布拉格公式,该现象证明了纤维素晶体中(200)平面的晶面间距有所降低[69]。类似的情况发生在碱性预处理中,碱液浸透结晶区,扰乱了纤维素晶体的有序堆砌,晶面间距增加,体系中的水分子进入双链晶胞单元,造成晶胞润胀,形成纤维素Ⅱ水合物,打开了纤维素Ⅱ的疏水平面,有利于酶吸附反应[15,70]。因此,通过一定的预处理方法扩展纤维素晶体疏水平面的晶面间距是提高酶解效率的有效方式。最新研究表明,随着酶解的进行,纤维素晶体的疏水晶面面积逐渐增大,且逐步发生自发聚集[71]。这一现象使得暴露在晶体外的羟基进一步减少,纤维素酶在纤维素晶体表面的吸附效率降低,难以进一步水解,直至酶解糖化达到终点。
上述预处理方法均可有效去除纤维素表面的生物基质(木质素与半纤维素),使得更多的微纤丝暴露出来,并在溶剂中发生润胀,增大纤丝与纤维素酶的接触面积。此时,纤维素链的氢键作用减弱,但在一定条件下可能发生共结晶和重结晶作用,造成结晶尺寸增加。Kuribayashi等[72]进一步研究了纤维素结晶尺寸变化和纤维素内部共结晶机理。研究发现,在水热处理温度超过180 ℃时,木材样品中纤维素结晶尺寸增加1倍,该结论与杨木液氨纤维爆破的结果相似[68]。高温打破了纤维间水分的结合,相邻纤维素晶胞之间亲水的(1-10)和(110)平面间距靠近,发生聚集重排,形成与纤维素链垂直的斜方晶系。值得注意的是,纤维素的这种共结晶现象增加了纤维素晶粒的疏水平面间距,使微纤丝进一步暴露,从而增加纤维素酶的接触位点,增效酶解反应。
![图1 植物细胞壁中纤维素多尺度超分子结构示意图[7]<br/>Fig.1 Scheme of aggregation of cellulose multi-scale supramolecular structure](2021年04期/pic16.jpg)
![图3 液氨处理纤维素Ⅰ转化为纤维素ⅢⅠ提高酶解效率示意图[27]<br/>Fig.3 NH3 pretreatment transformation of cellulose Ⅰ to cellulose ⅢⅠ and enhancement of enzymatic hydrolysis performance](2021年04期/pic18.jpg)
![图4 乙酰丙酸基低共熔溶剂处理毛竹原料小角X射线散射图[76]<br/>Fig.4 SAXS patterns for moso bamboo cellulose by levulinic acid based deep eutectic solvents pretreatments](2021年04期/pic20.jpg)
![图5 植物次生壁中纤维素、半纤维素(木聚糖为主)及木质素联接机制[79]<br/>Fig.5 A revised model of lignin-polysaccharide packing and secondary cell wall architecture](2021年04期/pic21.jpg)